Briología en el INECOL Dr. Efraín De Luna, Depto. Biodiversidad y Sistemática, INECOL, Xalapa, Ver |
Sinopsis de la clasificación de los musgos en Clases |
Relaciones filogenéticas
Las primeras proposiciones de relaciones filogenéticas dentro de los musgos interpretaron tres niveles fundamentales de organización estructural, considerando especialmente la morfología de las cápsulas. El linaje mas basal se consideró a las Sphagnopsida, con cápsulas operculadas. Las Andreaeopsida se colocaron como un grupo intermedio por sus cápsulas abriendo con valvas longitudinales. El grupo mas distal y diverso de los musgos verdaderos se clasificaron en las Bryopsida. Las cápsulas son operculadas y diferencian un peristoma. Los caracteres del exostoma y endostoma proveyeron la base para la clasificación de los grupos principales al nivel de orden y superiores.
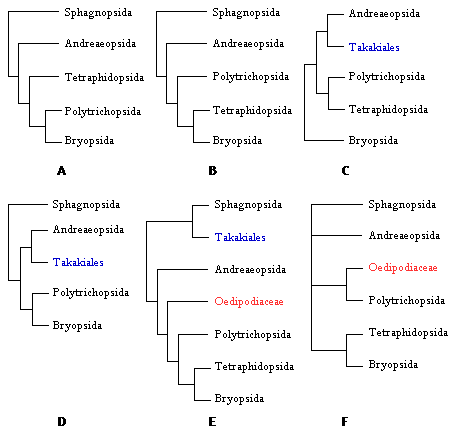
Las hipótesis modernas de las relaciones dentro de los musgos se basan en varios análisis cladísticos de datos morfológicos, ultraestructurales y moleculares. Un número reciente de la revista The Bryologist (Vol 103, numero 2, 2000) incluyo nuevos análisis filogenéticos que incorporan representantes de casi todas las familias de musgos. Cada contribución fue un gran esfuerzo colaborativo entre investigadores de varios países quienes conjuntaron datos de las secuencias de ADN de rbcL, rps4, trnL-F, y 18S rRNA (Fig. 28). | 5 Biología de los musgos 6 Filogenia y clasificación de los musgos Antecedentes históricos Sinopsis de las Clases |
Sintéticamente, Newton et al (2000) presentaron un análisis cladístico de todos los ordenes de musgos basado en datos morfológicos y cuatro juegos de secuencias del ADN para 33 representantes y 10 grupos externos. Los resultados incluyen la monofilesis de los musgos, la inclusión de Takakia como un musgo y hermano de Sphagnum y la monofilesis de los musgos con peristoma. En combinación, estos análisis recientes al nivel ordinal proveen el primer marco de referencia cladístico para los linajes principales de musgos. |
|
Clasificación
Actualmente se reconocen seis linajes principales, aunque su rango taxonómico como clases o subclases es controversial. En la mayoría de los casos, la clasificación a este nivel no esta totalmente establecida por que los mismos nombres se aplican a distintos grupos y rangos. Por ejemplo, "Bryopsida" se usa para el grupo que incluye todos los musgos si la división se denomina "Bryophyta", pero también se usa en sentido mas restringido para el grupo de los musgos con peristomas artrodontos. En este repaso, usamos nombres con el rango de clase para los cinco linajes principales.
El linaje mas basal es Sphagnopsida, hermano de un clado grande que incluye al resto de los musgos.
Entre los musgos con peristoma, el linaje mas basal es el grupo Polytrichopsida con peristomas nematodontos. Los dos linajes restantes tienen peristomas artrodontos. Tetraphidopsida es un clado pequeño, al igual que los tres anteriores. El quinto linaje, Bryopsida, incluye la gran mayoría de las especies de musgos. Los peristomas son haplolépidos y diplolépidos.
Es notable que la estructura de las relaciones entre Sphagnopsida, Andreaeopsida y los musgos con peristoma no ha cambiado sustancialmente. Este esquema básico de tres grandes grupos se ha expandido con la adición de grupos recientemente separados, como por ejemplo Ambuchania (Ambuchananiales) segregado de Sphagnum y Andreaeobryum (Andreaeobryales) relacionado con Andreaea.
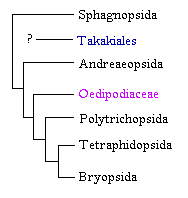
También se ha acomodado a Takakiales como grupo relacionado a los musgos sin peristoma. Inicialmente Takakia se clasificó entre las hepáticas del orden Calobryales. Con el descubrimiento de esporofitos y las secuencias de varios genes su lugar entre los musgos es difícil de refutar. No obstante aun es incierto si es el grupo hermano de Andreaeopsida o de Sphagnopsida.
Dentro del grupo de los musgos con peristoma, las ideas sobre las relaciones filogenéticas han oscilado entre dos panoramas especulativos. En uno, Polytrichopsida se consideraba como el grupo mas avanzado debido a la complejidad del gametofito (Brotherus 1925). En otros sistemas de clasificación, se interpreta este grupo como el mas ancestral debido a la simplicidad del peristoma nematodonto. Los análisis cladísticos modernos han corroborado la posición basal de los musgos con peristomas nematodontos en relación al origen mas reciente de los musgos con peristomas diplolépidos. No obstante, en estos análisis recientes persiste la incertidumbre sobre la posición de Tetraphidopsida y de la familia Oedipodiaceae.
|
División Bryophyta (musgos)
Clase 1 Sphagnopsida Clase 2 Takakiopsida Clase 3 Andreaeopsida Clase 4 Polytrichopsida Clase 5 Tetraphidopsida Clase 6 Bryopsida Subclase 1 Diphysciidae Subclase 2 Funariidae Subclase 3 Dicraniidae Subclase 4 Bryidae |
REFERENCIAS
Beckert S., Steinhauser S., Muhle H. y Knoop V.1999. A molecular phylogeny of the bryophytes based on nucleotide sequences of the mitochondrial nad5 gene. Pl. Syst. Evol. 218: 179–192. Cox C. J., Goffinet B., Newton A. E., Shaw A. J. y Hedderson T. A. J. 2000. Phylogenetic relationships among the diplolepideous-alternate mosses (Bryidae) inferred from nuclear and chloroplast DNA sequences. The Bryologist 103: 224–240. De Luna E., Buck W. R., Akiyama H., Arikawa T., Tsubota H., González D., Newton A. E. y Shaw A. J. 2000. Ordinal phylogeny within the hypnobryalean pleurocarpous mosses inferred from cladistic analyses of three chloroplast DNA sequence data sets: trnL-F, rps4, and rbcL. The Bryologist 103: 242–256. Goffinet B. y Cox C. J. 2000. Phylogenetic relationships among basalmost arthrodontous mosses with special emphasis on the evolutionary significance of the Funariineae. The Bryologist 103: 212–223. Hedderson T. A., Chapman R. L. y Rootes W. L.1996. Phylogenetic relationships of bryophytes inferred from nuclear encoded rRNA gene sequences. Pl. Syst. Evol. 200: 213–224. Hyvönen J., Hedderson T. A., Smith Merril G. L., Gibbins J. G. y Koskinen S. 1998. On phylogeny of the Polytrichales. The Bryologist 101: 489–504. La Farge C., Mishler B. D., Wheeler J. A., Wall D. P., Johanes K., Schaffer S. y Shaw A. J. 2000. Phylogenetic relationships within the haplolepideous mosses. The Bryologist 103:257-276. Mishler B. D. y Churchill S. P. 1984. A cladistic approach to the phylogeny of bryophytes. Brittonia 36: 406-424. Mishler B. D., Lewis L. A.,Buchheim M. A., Renzaglia K. S., Garbary D. J., Delwiche C. F., Zechman F. W., Kantz T. S. y Chapman R. L. 1994. Phylogenetic relationships of the ‘‘green algae’’ and ‘‘bryophytes.’’ Ann. Miss. Bot. Gard. 81: 451–483. Newton A. E., Cox C. J., Duckett J. G., Wheeler J., Goffinet B., Mishler B. D. y Hedderson T. A. J. 2000. Evolution of the major moss lineages. The Bryologist 103: 187–211. Shaw J. 2000. Phylogeny of the Sphagnopsida based on nuclear and chloroplast DNA sequences. The Bryologist 103: 277–306. | |
| |
x
x